|
(3)ヒメマス及びワカサギの生態
(1)ヒメマス
a. 形態
32尾の右第1鰓弓の鰓把数は27〜33本、14尾の幽門垂の数は40〜72本であった17)。
b. スモルト
2001年7月1日から7月16日にかけて、生出沖及び滝ノ沢沖10mに設置されたふくべ網に大量のスモルトが入網した。入網尾数は、生出では5日間で8,824尾、滝ノ沢では7日間で354尾であった。このうちの169尾(うち標識魚63尾)について魚体測定、年齢査定を行ったところ、被鱗体長は11〜18cm、モードは12cm、平均肥満度は11.7であった。年齢は、2年魚から4年魚で、組成は2年魚3%、3年魚91%、4年魚6%となっていた22)。
c. 移動分布
ア. 2001年7月5日にふ化場から放流された稚魚は、生出沖10mに設置されたふくべ網(放流場所から約300m)では、7月6日に11尾、7月7日に1尾採捕され、滝ノ沢沖10mに設置されたふくべ網(放流場所から約800m)では、7月10日から16日までの間に3尾が採捕されている。水深の浅い湖岸での採捕が少ないことから、放流稚魚は急速に沖合へ移動するものと考えられた22)。
イ. 水平垂直分布は、昼夜、水域、年級群により異なると思われる。垂直分布は7月において既に表層魚と深層魚の間に密度の上で著しい差が見られ、10月では更に明瞭になり、躍層を境にして両層にすみわける。中ノ湖では、中心に柱状に分布している魚群と周囲に点々と分布しているものがあり、中心のものは深度20〜150m付近まで点在した希薄な群を形成し、この群の中では40〜100mの間ではほぼ均等に分布している。周囲の沿岸近くに分布するものは躍層直下に最も多く分布し、下層に至るに従い逐次減少する。中ノ湖以外のところでは、中ノ湖の湖岸に近いものと類似した垂直分布密度を示している。中ノ湖の中心深部に分布するものは産卵親魚群であり、沿岸帯に近い躍層直下に分布しているものは未成魚群であると考えられている23)。
ウ. 躍層の浅い水域のヒメマスは遊泳深度が浅く、躍層の深い水域では遊泳深度が深い。産卵親魚も未成魚も夕刻には共に沿岸帯で釣獲されることから、昼間中ノ湖に分布していても夕刻になると浮上して沿岸一帯に分布するものと考えられ、毎日8km以上にわたる回遊をしていると考えられている23)。
エ. 水平分布については、7月の昼間では、沿岸帯で密度が濃く、沖合で薄い。沿岸部ではあっても中ノ湖のように急深の所は密度が濃い。10月では昼間には水深の深い所、特に中ノ湖の20m〜140mの水深に多くの魚が集まっており、湖の北半分にはきわめて密度が小さい。夜になると浮上し周囲に分散し、沿岸帯でもよく釣獲される23)。
オ. ヒメマスが夕方または朝方によく移動することは、タマズメまたは朝マズメとして知られている。7月、9月、10月では、夕刻から動きが活発になり、夜明けと共に離岸する傾向にあり、昼間は躍層近くに多く、水深50m付近にも分布するが、暗くなるに従って下層のものが浮上し始め、大部分が躍層直下の20m付近に集中する18)、19)、23)。
カ. 分布は水温と密接な関係があり、6月から10月では、7℃以下及び17℃以上では漁獲が激減する傾向にある。水深10mから20m付近で多く漁獲され、その水温帯は10℃〜15℃となっている19)。
キ. ふ化場のある西湖における回遊移動は、6月、7月では外部から回遊してくるものが多く、8月でははっきりした傾向は認められない。9月、10月では親魚の接岸移動が行われることもあり、漁獲が多くなる場合が多い。親魚では、産卵のため、早いものは6月から西湖へ回遊を始めるものもある18)、19)。
ク. 6月から10月の西湖には、時計周りと半時計周りに移動する2群があるものと考えられている20)。
ケ. ふ化場前で採捕した親魚に標識付けをして、1973年9月に金ケ森崎及び中山半島先端から、1974年10月には子ノ口及び滝ノ沢から、一カ所当たり15尾〜30尾を放流したところ、早いものは翌日、遅くても2週間以内にふ化場前に回帰することが確認されている。再捕率は1973年には87.5%、1974年には48.3%となっている19)。
d. 天然産卵
ア. 1967年から1977年に行われた調査で天然産卵床が確認されている。天然産卵床は、水深0.5mから最も深い所で8mにかけて分布し、1〜3mの水深範囲に多い。その大きさは、0.2m2から0.35m2と小さく発見しにくい。産卵床は一つ一つ独立して存在することは珍しくほとんど群床を形成し、時には床が重複し合って一大産卵場を形成する。1976年には中山半島西側湖岸一帯に多数の産卵床からなる産卵場が形成され、1979年には湖内で81箇所の産卵床が確認されている。底質は、1973年では径3〜5cmの小石、1975年、1976年では礫、砂となっている18)、19)、21)。
イ. 天然産卵は、表面水温が13℃〜14℃になる10月下旬頃から8℃〜9℃になる11月上旬に行われるものと考えられる19)。
e. 漁獲変動及び成長
ヒメマス資源は、1980年代前半まで比較的高水準で安定した状態にあった。しかし、1985年にワカサギが大量に漁獲され、ヒメマス漁獲量が著しく減少してから、両種は大きな資源変動を繰り返すようになっている。
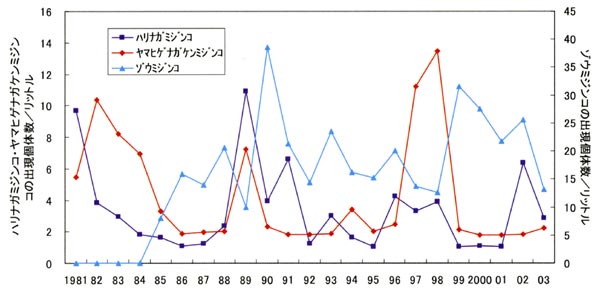
ア. ヒメマスの餌であるヤマヒゲナガケンミジンコ、ハリナガミジンコが多く出現した時期は、ヒメマス漁獲量の多い時期と一致し、両プランクトンの年平均密度とヒメマス漁獲量には正の相関が認められ、ヒメマス漁獲量と両プランクトンが連動して変化すると考えられている31)。
イ. ヒメマス年級群の平均漁獲年齢は、ヒメマス若齢期のハリナガミジンコ、ヤマヒゲナガケンミジンコの年平均密度と負の相関を示し、ヒメマス年級群の生残率と、放流された年に発生したワカサギ年級群の豊度には負の相関関係が認められ、ヒメマスとワカサギは同じ餌をめぐって強い競争関係にあると考えられている26)、27)、28)。
ウ. 漁獲されたヒメマスの年齢は、標識と鱗相から判断すると、2年魚から7年魚で、各年魚及び主体となる年魚の割合は、漁獲年により大きく異なる。例えば、1991年では2年魚84%、1992年では3年魚94%、1995年では4年魚85%、1997年から1999年では3年魚が60〜67%を占めており、平均すると各年の漁獲主体は、多い順に3年魚、4年魚、2年魚となっている1)。
エ. ヒメマスの大きさも漁獲年によって大きく異なり、平均的な成長パターンを求めることはできない。内臓を除去して集荷場へ水揚げされたヒメマスの平均体重について最大と最小を示すと、3年魚では49g(1987年)〜160g(1991年)、4年魚では73g(1988年)〜209g(1999年)となっている1)。
(2)ワカサギ
沢目ほか22)、長崎・沢目11)、水谷14)、鈴木28)などに基づき、十和田湖におけるワカサギの生態の概要について記載する。
a. 産卵生態
ア. 産卵時期は4〜7月で、盛期は年及び場所により異なり、1983年は5月中旬、2003年は7月(河川水温20℃前後)であった。
イ. 産卵は、銀山沢、大川沢、鉛沢、宇樽部沢などの流入河川河口付近で行われ、水深10〜30cmの砂礫底に産着卵が確認できる。
ウ. 産卵期にはオスの割合が高く、産卵親魚は、大型であるほど早期に成熟、産卵する。
エ. 浮遊仔魚の出現状況から、産卵ピークは少なくとも2回あると推定される。
b. 成長
ア. 生出から発荷地先100m付近で行った2003年の調査では、7月10日に脊索長4〜11mm、8月8日に24〜11.1mmの仔魚が採捕されている。
イ. 1998年8月には、湖岸から沖合10m程度の地点で全長2〜3cmのワカサギ稚魚が大量に群泳しているのが目視されている。稚魚は、鉛沢、ふ化場前、鉛山などでも確認されており、湖内全域に分布していると推察された。
ウ. 春にふ化したものは、翌年春に0歳で産卵に参加する。その翌年の1歳魚及びさらに翌年の2歳魚も、それぞれ産卵に参加することが確認されている。
エ. 成長は年により大きな変動があり、1983〜1997年における1歳魚親魚の体長は6.1〜9.5cmであった。この変動は、ヤマヒゲナガケンミジンコの密度及び降水量と正の相関が認められている。
オ. 十和田湖において記録が残っている最大個体は、1984年5月5日に神田川で採捕された体長144mm、体重41.8gの個体である。
c. 食性
ア. 1998年2〜10月に採捕されたワカサギの胃内容物は、時期やサイズにより異なっていた。8月における体長24〜27mmの稚魚は、最も個体数が多いのはハリナガミジンコで、ヤマヒゲナガケンミジンコがそれよりやや少なく、次いでカイアシ類幼生であった。その他、枝角類、カイアシ類、ユスリカ類の幼虫や蛹なども確認したが少なかった。また、全長5〜9mmのヌマチチブの仔魚と推定される魚類を摂餌していたものも認められた。
イ. 未成魚、成魚は、ハリナガミジンコとヤマヒゲナガケンミジンコを主体に、ゾウミジンコ、枝角類、ユスリカ類の幼虫や蛹の他、ウキゴリ仔魚なども出現した。
ウ. 1999年5〜10月に採捕されたワカサギの胃内容物として認められたものは、ゾウミジンコを主体に、カイアシ類、シカクミジンコ属、ユスリカの幼虫、蛹及び成虫、陸生昆虫、魚類、ハリナガミジンコなどであった。
d. 被食
ア. ヒメマス:体重30g未満のものから各サイズのものが、ワカサギを摂餌していた。ワカサギを飽食していた個体も認められたが、全体としてはヒメマスのワカサギに対する餌料としての価値は大きくないと推察される。
イ. サクラマス:体長20cm程度の個体が体長3〜4cmのワカサギ2個体を摂餌するなど、ワカサギの摂餌が散見される。
ウ. イワナ:体長26cm、体重279gの個体がワカサギ2.8gを摂餌していた。
エ. ヌマチチブ:6月のワカサギ産卵期に、ワカサギ卵が認められた。
秋田県水産振興センターでは、ヒメマスの主な餌の出現状況を把握するため、湖内の10箇所において年3回(6、8、10月)のプランクトン調査を実施している。また、ヒメマスやワカサギなどが何を餌としているのかを把握するため、青森県水産総合研究センター内水面研究所が刺し網で採捕した魚類の胃内容物調査もあわせて実施している。
これらの調査結果から、1993年10月、1996年10月、1997年10月、1998年8月、2001年10月のようにハリナガミジンコの出現水準が高い時期には、ヒメマスは大きさにかかわらずハリナガミジンコを好んで捕食している様子がうかがえる。1980年代以前には認められていたように、ヤマヒゲナガケンミジンコもヒメマスに選択的に捕食される傾向はうかがえるが、近年は大量に出現することがほとんどないため、ハリナガミジンコほど目立っ存在ではなくなってしまった。一方、同じ枝角類でもより小型のゾウミジンコについては、大量に出現していても捕食しているヒメマスはほとんど認められない。
なお、低い水準が続いた後ハリナガミジンコの出現水準が上昇すると、湖内のヒメマスが急激に体長を増し肥満度も高くなり、それに伴って漁獲量も気に好転するという状況がしばしば認められる。その顕著な例が1997年から1998年にかけての状況で、1997年春から1998年秋にヤマヒゲナガケンミジンコが、1997年秋から1998年秋にハリナガミジンコが例年になく高水準で連続して出現したことにより、1995年、1996年と続いた不漁から1997年には15.5tと6年ぶりに漁獲量が10tを超え、翌1998年には43tの漁獲量を記録するとともに、秋には過去最高と思われる85,000尾を超える親魚が十和田湖増殖漁業協同組合ヒメマスふ化場前に接岸し、ふ化場から湖へ続く水路がヒメマスで埋め尽くされるといった状況が確認された。
ハリナガミジンコが少ない時には、小・中型のヒメマスは陸生昆虫を、中・大型のヒメマスはワカサギなどの魚類やヨコエビ類を捕食している場合が多くなるが、このような傾向が続く時期にはヒメマスの肥満度が低く漁獲量も少ないことからも、ハリナガミジンコの餌料としての重要性が示唆される。ただし、ヨコエビ類については大型のヒメマスほど好む傾向が強いようで、ハリナガミジンコが比較的多いときでもこれが大量に捕食されていることがある。
このように、ハリナガミジンコなど大型動物プランクトンに対する依存度が極めて高いことが推察されるヒメマスではあるが、1998年の春にはハリナガミジンコの出現水準が高かったにもかかわらず小型から大型に至るあらゆるサイズのヒメマスの胃内容物としてユスリカ類が優占していた。ユスリカ類がヒメマスやワカサギ、サクラマスの幼魚などに捕食されるのは主としてさなぎのステージで、春から夏にかけてこれらの魚類の胃内容物として優占する場合があるが、このときの状況から大量に羽化する時期にはヒメマスはハリナガミジンコよりもユスリカ類を選択的に捕食する可能性があることも示唆された。
ワカサギもヒメマスと同様に代表的なプランクトン食魚とされており、やはりハリナガミジンコの出現水準が高い時期にはほとんどの個体がこれを餌としている。ただし、ヒメマスとは違って、ハリナガミジンコが少ない場合でもゾウミジンコが多いときにはこちらを捕食している個体も多い。従って、ワカサギはハリナガミジンコを選択する傾向が認められるものの、ゾウミジンコについても季節を問わず餌料として利用していると考えられる。これらの他には陸生昆虫、ユスリカ類のさなぎや幼虫、ハゼ科魚類などの稚・仔魚などを捕食している個体もいるが、プランクトンに比べると利用頻度は高くない。
そのほか、サクラマス、イワナといった大型に成長するサケ科魚類は小型の時には水生昆虫や陸生昆虫を、大型魚ではワカサギを主体とした魚類を餌とし、ウキゴリ、ヌマチチブなどのハゼ科魚類は水生昆虫などの底生生物や魚類の稚仔魚を、イトヨやイバラトミヨといったトゲウオ類は底生生物、ワカサギなどの卵、動物プランクトンなど様々なものを、コイ、ギンブナは湖底の底生生物や有機物などを、それぞれ主な餌料として利用している。
| 注)同年の6、8、10月調査分の平均値をその年の値とした |
外来種(alien species)とは、過去あるいは現在の自然分布域外に導入(人為によって直接的・間接的に自然分布域外に移動させること)された種、亜種、あるいはそれ以下の分類群を指し、生存し繁殖することができるあらゆる器官、配偶子、種子、卵、無性的繁殖子を含むものをいい、外来種はその起源によって国外外来種と国内外来種に分けられる。
近年・外来種問題が大きく取り上げられるようになってきたのは、「国外または国内の他地域から・野生生物の本来の移動能力を越えて、人為によって意図的・非意図的に導入された種である移入種(外来種)が、地域固有の生物相や生態系に対する大きな脅威」10)となってきているからである。
これら移入種(外来種)は、魚類においては「外来魚」と呼ばれ、特に、オオクチバス、ブルーギル、コクチバスなどの外国産外来魚は、在来魚に対する影響や漁業被害と関連して取り上げられる機会が多い。
このような観点から、現在、十和田湖に生息している魚類について考えてみよう。
結論から言えば、十和田湖に生息している魚種はすべて外来魚ということになる。すなわち、魚類が生息していなかった十和田湖に現在生息が認められている魚種は、すべて意図的あるいは非意図的に人間が移動させたものであると推察される。
しかし、これら外来魚の導入の目的、経緯や時期について不明なものが多いことも特徴の一つとしてあげられる。また、導入(放流)後、湖内で天然繁殖しなかったと考えられる魚種も少なくない。さらに、最近になってオオクチバスやブラウントラウトなど、他水域で在来魚に大きな影響を与えているとして駆除などが行われている魚種が湖内で確認され、地元では危機感を募らせているのも事実である。
一方、ここで留意しなくてはいけないことは、外来魚だからすべて問題であるということではなく、ヒメマスのように地元の漁業や観光などの地場産業と密接な関わりを持ち、地元にとってきわめて重要な魚種も存在しているということである。
現在、十和田湖に生息しているヒメマスは、ほぼそのすべてが十和田湖増殖漁協による増殖(人工種苗生産、放流)に由来している。すなわち、魚類が生息していなかった十和田湖において意図的にヒメマスを導入し、漁業権を有してヒメマスを管理している同漁協にとって、ヒメマスの生息に影響を与える(与える可能性のある)魚種は排除の対象となるのは当然のことである。
実際、ワカサギは同漁協の漁業権の内容魚種とはなっていないが、ヒメマス資源に大きな影響を与えることから積極的に駆除を行っている。同様に、オオクチバス、コクチバス、ブルーギルなどヒメマス資源に影響を与える可能性のある魚種については、今後とも湖内に放流されないよう注意する必要がある。
1)青森県内水面研究所内部資料
2)秋田県水産試験場. 十和田湖鱒孵化ニ関スル調査. 明治39年度秋田県水産試験場報告, 1906; 43-51.
3)秋田県水産試験場. 鮭鱒増殖事業. 昭和3年度試験事業報告, 1930; 90-109.
4)秋田県水産試験場. 鮭鱒増殖事業. 昭和4年度試験事業報告, 1931; 87-102.
5)天野勝三. 十和田湖の水温とヒメマス・ワカサギの漁獲変動. 平成13年度事業報告書, 青森県内水面水産試験場, 2003; 35-41.
6)原子保. 青森県におけるサケ科魚類相. 青森県水産試験場研究報告, 2002; 23-28.
7)Higuchi M & Goto A. Genetic evidence supporting the existence of two distinct species in the genus Gasterosteus around Japan. Environmental Biology of Fisheries 1996; 47: 1-16.
8)疋田豊彦・谷口定利. 十和田湖に棲息している魚類. 北海道さけ・ますふ化場研究報告, 1959; 13: 45-48.
9)細谷和海. サケ科「日本産魚類検索−全種の同定−」(中坊徹次編). 東海大学出版会, 東京, 1993; 256-261.
10)環境省野生生物保護対策研究会移入種問題分科会. 移入種(外来種)への対応方針, 2002.
11)長崎勝康, 沢目司. 十和田湖資源対策調査. 平成9年度事業報告書, 青森県内水面水産試験場, 1999; 26-35.
12)丸山為蔵, 藤井則一, 木島利通, 前田弘也. 外国産新魚種の導入経過. 水産庁研究部資源課・水産庁養殖研究所, 1987; 157pp.
13)宮地伝三郎, 川那部浩哉, 水野信彦. 原色日本淡水魚類図鑑. 保育社, 大阪, 1976; 462pp.
14)水谷寿. 十和田湖の生態系および資源対策調査結果(1998年). 国立環境研究所研究報告, 1999; 146: 137-150.
15)水谷寿. 十和田湖資源対策調査. 平成12年度秋田県水産振興センター事業報告書, 1999; 242. 267.
16)森誠一. 十和田湖沿岸域の魚類, 特にイトヨの生態を中心に. 国立環境研究所研究報告, 1999; 146: 95-109.
17)頼茂. ヒメマス天然産卵調査. 昭和44年度十和田湖資源対策事業調査報告書, 十和田湖ふ化場協議会1960; 1-19.
18)頼茂. 昭和48年度十和田湖資源対策事業調査報告書. 十和田湖ふ化場協議会, 1974.
19)頼茂. 資源調査. 十和田湖資源対策事業調査報告書(昭和42〜55年度調査結果の総括), 十和田湖ふ化場協議会, 1981; 1-53.
20)頼茂, 松田銀治, 岩谷良作. 昭和47年度十和田湖資源対策事業調査報告書. 十和田湖ふ化場協議会, 1973.
21)頼茂, 金沢広重, 米谷峯夫. 資源調査. 昭和50・51年度十和田湖資源対策事業調査報告書, 十和田湖ふ化場協議会, 1977; 1-33.
22)沢目司, 天野勝三, 田村直明, 松田忍. 十和田湖資源対策調査. 平成13年度事業報告書, 青森県内水面水産試験場, 2003; 16-34.
23)白石芳一. 魚群探知機による十和田湖の魚族資源調査. 昭和44年度十和田湖資源対策事業調査報告書, 十和田湖ふ化場協議会, 1960; 1-9.
24)杉山秀樹. 秋田の淡水魚. 秋田魁新報社, 1985; 168pp.
25)杉山秀樹. 田沢湖まぼろしの魚クニマス百科. 秋田魁新報社, 2000; 239pp.
26)水産庁さけ・ます資源管理センター. 十和田湖におけるヒメマスとワカサギの成長. 平成11年度十和田湖水質・生態系資源対策会議資料 2000
27)鈴木俊哉, 長崎勝康, 水谷寿, 帰山雅秀. 1999. 十和田湖におけるヒメマスおよびワカサギの個体群動態. 国立環境研究所研究報告 146, 27-35.
28)鈴木俊哉. 十和田湖におけるヒメマスとワカサギの資源変動要因. 平成14年度十和田湖資源対策会議資料, 2002.
29)高田啓介. トミヨ属魚類の遺伝学的分化「日本の淡水魚類−変異・分布・種分化をめぐって」(水野信彦, 後藤晃編)東海大学出版会, 東京, 1987; 135-143.
30)高村健二. 十和田湖イトヨの由来の系統的解析. 国立環境研究所研究報告, 2001; 167: 102. 105.
31)高村典子, 三上一, 水谷寿, 長崎勝康. ワカサギの導入に伴う十和田湖の生態系の変化について. 国立環境研究所研究報告, 1999;146: 1-15.
32)竹内基, 松宮隆志, 佐原雄二, 小川隆, 太田隆. 青森県の淡水魚類相について. 淡水魚1985; 11: 117-133.
33)徳井利信. 十和田湖の湖水型サクラマス(Oncorhynchus masou)について. 水産増殖1962; 10: 133-136.
34)徳井利信. 十和田湖漁業史. 徳井淡水漁業研究所, 1984; 233pp.
35)十和田湖ふ化場協議会. 十和田湖資源対策調査, 1982; 125pp.
|