|
3、細胞内サイトカインの解析
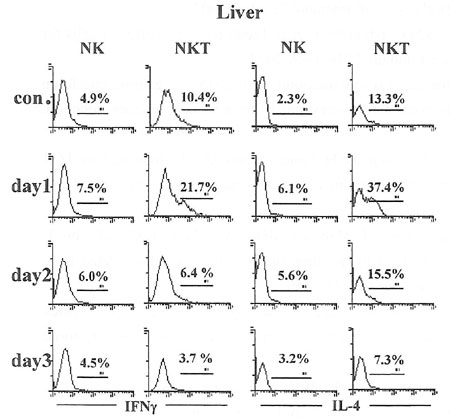
細胞内サイトカインの解析については肝臓NK、NKT細胞は絶食1日目でIFN-γ及びIL-4が増加したものの、2、3日目では減少を認めた(図5)。脾臓は肝臓と同様な変化であった(data not shown)。
|
図5.
|
肝臓細胞内内サイトカインの解析を行った。図内の数値は各分画細胞のサイトカインの比率を示す。
|
4、絶食マウス肝臓及び脾臓リンパ球のNK細胞傷害活性の変化
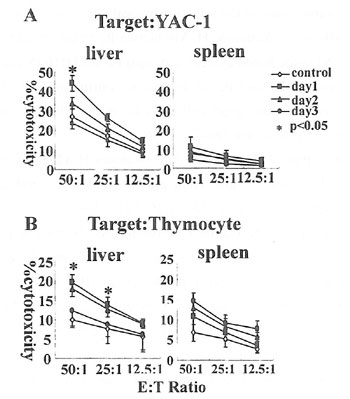
絶食マウスNK細胞傷害活性の変化について、YAC-1及びB6マウス胸腺細胞を標的細胞として、絶食後1、2、3日目で、肝臓及び脾臓リンパ球のNK細胞傷害活性を測定して、NK細胞比率あたりの細胞傷害活性を解析した(図6)。肝臓では絶食1日目でYAC-1(図6A)及びB6胸腺細胞(図6B)にて、細胞傷害活性が最大となり、2、3日目では徐々に低下した。一方、脾臓では、YAC-1及びB6マウス胸腺細胞に対する細胞傷害活性が絶食の前後で著明な差を認めなかった。
|
図6.
|
図6. 肝臓及び脾臓内単核球をエフェクターとした。標的細胞は(A)YAC-1(B)B6胸腺細胞を用いた。
|
考察
今回の研究は、絶食マウスのリンパ球による性質と機能の変化について解析した。対照組より、短期絶食マウス肝内NK細胞は、強い細胞傷害活性を有することが明らかになった。絶食により、胸腺の萎縮と、胸腺外分化T細胞であるIL-2Rβ+CD3int細胞15)の増加がみられたことから、自然免疫系が活性化されたことによると考えられる。
われわれは、以前の研究において、ストレスが自然免疫系を活性化させ、逆に獲得免疫系の働きを弱めることを報告してきた16),17)。本研究により、絶食マウスも、このストレス反応に似た反応を示すことがわかった。ストレスは胸腺の萎縮を誘導し、通常のT細胞の分化を抑制し、獲得免疫系を不活性化し、逆に自然免疫系を活性化する18)。ただし、24時間の短期絶食は自然免疫を担うNK、NKT細胞の機能増強を誘導し、自然免疫系を活性化させるとはいえ、それ以上になると細胞数、機能とも低下することが明らかとなった。
従来、絶食は免疫応答低下を引き起こすといわれてきたが、本研究により、絶食時間によっては免疫応答の中でも自然免疫を増強しうることが示唆された。今後、自然免疫を増強する因子及び粘膜免疫を担う小腸のCD8αα+B220+γTCR+細胞についても検討する予定である。
参考文献
1. Weerasinghe A, Sekikawa H, Watanabe H, Mannoor MK, Morshed SRM, Halder RC, Kawamura T, Kosaka T, Miyaji C, Kawamura H, Seki S and Abo T. Association of intermediate T cell receptor cells, mainly their NK 1.1- subset, with protection from malaria. Cell Immunol 207: 28-35, 2001.
2. Abo T, Kawamura T and Watanabe H. Physiological responses of extrathymic T cells in the liver. Immunol Rev 174: 135-149, 2000.
3. Mannor MK, Weerasinghe A, Halder RC, Morshed SRM, Ariyashinghe A, Watanabe H, Sekikawa H and Abo T. Resistance to malarial infection is achieved by the cooperation of NK1.1+ and NK1.1- subsets of intermediate TCR cells which are constituents of the innate immunity. Cell Immunol 211: 96-104, 2001.
4. Mannor MK, Halder RC, Morshed SRM, Ariyashinghe A, Bakir HY, Kawamura H, Watanabe H, Sekikawa H and Abo T. Essential role of extrathymic T cells in protection against malaria. J Immunol 169: 301-306, 2002.
5. Changchun Li, Xuefeng Bai, Sen Wang, Chikako Tomiyama-Miyaji, Toru Nagura, Toshihiko Kawamura and Toru Abo. Immunopotentiation of NKT cells by low-protein diet and the suppressive effect on tumor metastasis. Cellular Immunology 231: 96-102, 2004.
6. Kawamura H, Kameyama H, Kosaka T, Kuwahara O, Bannai M, Kawamura T, et al. Association of CD8+ natural killer T cells in the liver with neonatal tolerance phenomenon. Transplantation 73: 978-94, 2002.
7. Ishimoto Y, Tomiyama-Miyaji C, Watanabe H, Yokoyama H, Ebe K, Tsubata S, et al. Age-dependent variation in the proportion and number of intestinal lymphocyte subsets, especially natural killer T cells, double-positive CD4+ CD8+ cells and B220+ T cells, in mice. Immunology 113: 371-7, 2004.
8. Takahashi S, Kawamura T, Kanda Y, Taniguchi T, Nishizawa T, Iiai T, et al. Activation of CD1d-independent NK1.1+ T cells in the large intestine by Latobacilli. Immunol Lett 102: 74-8, 2006.
9. Bakir HY, Tomiyama-Miyaji C, Watanabe H, Nagura T, Kawamura T, Sekikawa H, et al. Reasons why DBA/2 mice are resistant to malarial infection: expansion of CD3intB220+γδT cells with double-negative CD4-8- phenotype in the liver. Immunology 17: 127-35, 2006.
10. Yamagiwa S, Sugahara S, Shimizu T, Iwanaga T, Yoshida Y, Honda S, et al. The primary site of CD4-8-B220+ T cells in lpr mice: the appendix in normal mice. J Immunol 160: 2665-74, 1998.
11. Tsukada C, Yokoyama H, Miyaji C, Ishimoto Y, Kawamura H, Abo T. Immunopotentiation of intraepithelial lymphocytes in the intestine by oral administrations of β-glucan. Cell Immunol 221: 1-5, 2003.
12. Miyaji C, Watanabe H, Miyakawa R, Yokoyama H, Tsukada C, Ishimoto Y, et al. Identification of effector cells for TNFα-mediated cytotoxicity against WEHI164S cells. Cell Immunol 216: 43-9, 2002.
13. Emoto M, Neuhaus O, Emoto Y, Kaufmann SHE. Influence of β2-microglobulin expression on gamma interferon secretion and target cell lysis by intraepithelial lymphocytes during intestinal Listeria monocytogenes infection. Infect Immun 64: 569-75, 1996.
14. Watanabe T, Kawamura T, Kawamura H, Haga M, Shirai K, Watanabe H, Eguchi S and Abo T. Intermediate TCR cells in mouse lung. Their effector function to induce pneumonitis in mice with autoimmune-like graft-versus-host disease. J Immunol 158: 5805-5814, 1997.
15. Halder RC, Kawamura T, Bannai M, Watanabe H, Kawamura H, Mannoor MDK, Morshed SRM and Abo T. Intensive generation of NK1.1- extrathymic T cells in the liver by injection of bone marrow cells isolated from mice with a mutation of polymorphic MHC antigens. Immunology 102: 450-459, 2001.
16. Murayama S. Tsukahara A, Suzuki S, Tada T, Minagawa M, Watanabe H, Hatakeyama K and Abo T. Quick recovery in the generation of self-reactive CD4low NKT cells by an alternative intrathymic pathway when restored from acute thymic atrophy. Clin Exp Immunol 117: 587-595, 1999.
17. Oya H, Kawamura T, Shimizu T, Bannai M, Kawamura H, Minagawa M, Watanabe H, Hatakeyama K and Abo T. The differential effect of stress on NKT and NK cell function. Clin Exp Immunol 121: 384-390, 2000.
18. Tagoh H, Nishijo H, Uwano T, Kishi H, Ono T and Muraguchi A. Reciprocal IL-1 beta gene expression in medial and lateral hypothalamic areas in SART -stressed mice. Neurosci Letters 184: 17-20, 1995.
|