|
4. 論議
ハナサキガニの「根室半島−四島水域」個体群とサハリン個体群との間に、遺伝的差異が存在するという事実は、これらの個体群の浮遊幼生が輸送分散される領域を、確実に隔離する機構が存在することを意味する。しかし、モデルによる実験では千島海盆の低気圧性循環により、サハリンからの粒子が根室半島−四島水域に到達しており、隔離機構は存在しない。モデルでは東サハリン海流は再現されていたが、千島海盆の流れが異なり、宗谷暖流も十分再現されていない。このことは、逆の意味で隔離機構について、ひとつの仮説;宗谷暖流が幼生の輸送を隔離している可能性、を示唆している。
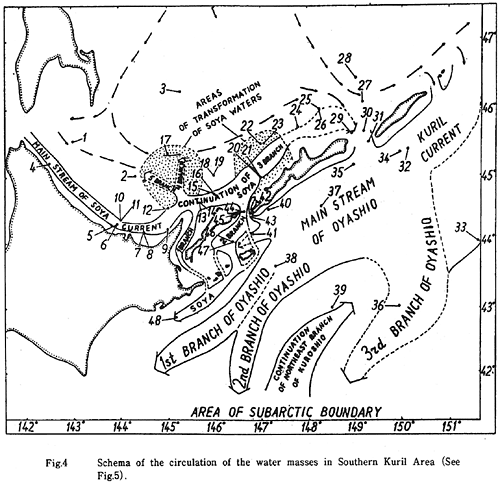
Bobkov(1993)の南千島周辺海域の流れの模式図(図28)によれば、北海道のオホーツク海沿岸から知床半島と国後島のオホーツク海側、さらには択捉島のオホーツク海側は、宗谷暖流の主流によって、東サハリン海流の流れから切り離されている。
したがって、宗谷暖流が、ハナサキガニの根室半島−四島水域個体群とサハリン個体群との、遺伝的隔離をもたらしている、という作業仮説が浮かび上がる。
図28. 南千島周辺海域の流れの模式図(Bobkov, 1993).
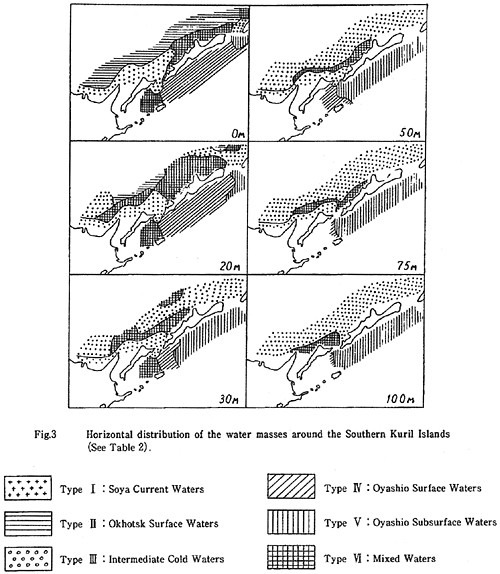
もしハナサキガニの浮遊幼生が環境選択を行うとすれば、水塊の違いも重要な要素である。Bobkov(1993)は南千島周辺海域の水塊分布の模式図も示している(図29)。これによれば、宗谷暖流水(Type I: 水温14-17℃)が北海道沿岸から知床半島、そして国後島のオホーツク海側の75m深までを覆っており、東サハリン海流の水塊を含むオホーツク海表層水(Type II: 水温8-14℃)はその外側の表層(0m)に限られている。オホーツク海表層水の下部(20m以深)には非常に冷たい中冷水(Type III: 水温1.0-2.2℃)が分布している。
したがって、ハナサキガニの浮遊幼生が、オホーツク海表層水の水温帯(水温8-14℃)を適水温とし、20m以浅の浅い岩礁域に着底するように能動的な環境選択をするものとすれば、宗谷暖流水の存在は、東サハリン海流の水塊から四島水域に到達することを、遮断することになるだろう。
今回はグローコテ期の行動を考慮していない。根室市水産研究所による飼育実験では、ゾエア期・グローコテ期とも21日であり、グローコテ期に着底場所を探索することがわかっている(Kittaka、2003)が、グローコテがどのくらい自由に泳げるのか、どのような条件で着底場所を決めるのかなどは分かっていない。もし今回の設定より浮遊期間が短いのだとすれば、サハリン周辺起源の浮遊幼生が北海道沿岸にたどり着く可能性はさらに低くなると考えられる。
図29. Bobkov(1993)の南千島周辺海域の水塊分布の模式図
親潮と宗谷暖流は、このモデルでは十分再現できなかった。宗谷暖流は、サハリン起源の幼生が四島海域に到達することを妨げる主要な要因である可能性があり、その再現は、きわめて重要である。親潮は、オホーツク海全体の流れに影響するとともに、北方四島周辺の浮遊幼生の輸送に大きな影響を与えることが予想される。
ハナサキガニのサハリン系個体群から根室半島−四島海域に浮遊幼生が補給されないということは、サハリン沿岸を経由する(海面下に浮遊する)汚染物質も、この海域には到達しにくいことを意味する。
ハナサキガニの浮遊幼生がふ化する時期は,オホーツク海の流氷が融けて後退する時期であり、幼生が浮遊期を過ごすのはごく浅いオホーツク海表層水であり、その直下は生存を脅かす可能性のある冷たいオホーツク海中冷水が存在している。したがって、ハナサキガニ幼生の生活環境であるこれらの海洋構造とその出現は、流氷と冬から春への季節の節目に支配されており、気候変動の影響を受けて大きく変動する可能性を持っている。
根室半島−四島海域の個体群は、ハナサキガニの分布域の南限に位置する。しかも、宗谷暖流が北方のサハリン沿岸の個体群からの浮遊幼生の供給を遮断しているとすると、温暖化が根室半島−四島海域のハナサキガニ個体群に大いに影響することが懸念される。
これらの問題に答えるためには、以下の課題を解決してこの研究を発展させることが必要である。
1. 宗谷暖流と親潮が再現されるように境界条件を改良する。
2. ランダムウォークによる拡散項をいれる。
3. 地形をより現実的に改良する。
4. 三角水域周辺の部分モデルとオホーツク海全体のモデルとを結合し、流れの再現性を改善する。
5. モデルに潮汐を組み込み、meso-scale eddyと千島列島周辺の強い潮汐混合の再現を試みる。
6. 海岸線にいる粒子の挙動がより現実に近くになるように改善する。
7. 水温場が再現性を確認し、水温に対する幼生の行動を考慮して、幼生の分散過程を調べる。
8. 粒子追跡モデルを三次元に変更し、幼生の鉛直移動を考慮する。
9. 低次生産系生態系モデルを組み込んで、ゾエア期幼生の摂餌と生き残り・成長の影響を調べる。
謝辞
この研究は、根室市ハナサキ・プログラムの一環として、日本財団の助成金を受けて実施されたので、ここに記して謝意とする。また、データの使用を快諾いただいた北海道大学低温科学研究所大島慶一郎助教授に深く感謝する。
引用文献
Bobkov, Andrew A. 1993: Umi to Sora, Vol.69, No.1, pp.41-52.
Kittaka, J., 2003: Biological and ecological characteristics of Paralithodes brevipes in coastal water of Nemuro. Proceedings of the 1st Hanasaki Program Workshop, Mar., 2003, Nemuro, Hokkaido.
Nakamura, T., T.Awaji, T.Hatayama and T.Akimoto, 2000: Tidal exchange through the Kuril Straits, J.Phys. Oceanogr., 30, 1622-1644.
Ohshima, K.I., M.Wakatsuchi, Y.Fukamachi and G.Mizuta, 2002: Near-surface circulation and tidal currents of the Okhotsk Sea observed with satellite-tracked drifters. J.Geophys. Res., 107, 3195, doi: 10. 1029/2001JC00I005.
Ooshima, K.I., Y.Fukamachi, T.Mutoh and M.Wakatsuchi, 2005: A generation mechanism for mesoscale eddies in the Kuril Basin of the Okhotsk Sea: Baroclinic instability caused by enhanced tidal mixing. J.Oceanogr, Vol.61, pp.247-260.
Ikeda, M., H.Uyama and N.Taniguchi, 2005: Genetic diversity and population structure of Hanasaki crab: Aspects from DNA analyses. Proceedings of the 3rd Hanasaki Program Workshop, Mar.2 - 5, 2005, Nemuro, Hokkaido.
Torisawa, M., S.Kohno. K.Sakamoto and I.Hakata, 1999: Growth in the early life stage of the spiny king crab, Paralithodes brevipes. Sci. Rep. Hokkaido Fish. Exp. Stn., (55):161-167.
|