|
結果と考察
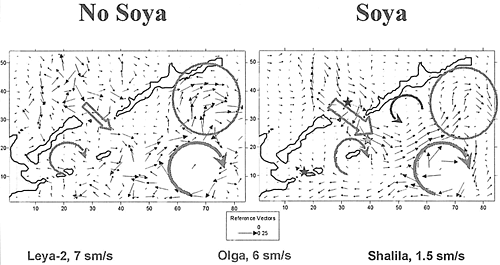
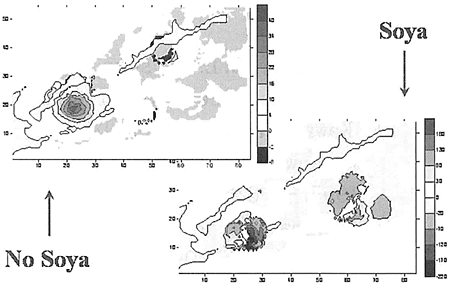
海面の流れの場のモデリングにおいて外部からの擾乱がないものとすると、モデルの南東部の範囲に時計回りの渦が見られる(毎秒40cm以下)。この渦は、択捉島の南の沿岸にかかる流れを起こしている(毎秒30cm以下)。オホーツク海側では流れの場に安定した構造は見られない。太平洋とオホーツク海の間の水交換は、国後水道を通じて行われていることに注目できる(毎秒10cm以下)。南千島海峡の中央部には、時計回りのぼんやりと現れた渦を見ることができる(毎秒8cm以下)。太平洋側の海水が、南千島列島の島々の間から南千島海峡に流入していることが見られる(毎秒5cm以下)。
宗谷海流は、北海道より北のモデルの範囲のセルの一部に速度を追加してやって(毎秒18cm以下)入れた。海水面の流れの場は、次のように形が変わった。オホーツク海側では、流れの構造が一層秩序だったものになった。国後水道を通るオホーツク海の海水の水流が強くなった(毎秒15cm以下)。この流れと太平洋の渦の相互作用によって部分的に渦が弱まるが、それはモデルの南部の範囲で海流に反映された。同時に択捉島より南の流れは弱められた(毎秒10cm以下)。択捉島より南にある渦(反時計回り)と色丹島の周りにある渦(時計回り)の二つが、一層はっきり現れるようになった。
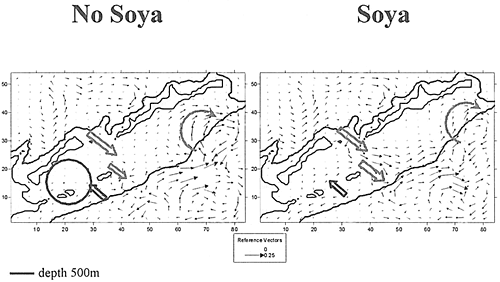
底層の流れの場は、深海部における擾乱がないとすると大陸棚の辺縁部のラインにつながっている。択捉島の南の部分にかかっている時計回りの渦(毎秒10cm以下)の分岐が保たれていることに注目したい。国後水道における水の方向と動きは保たれている(毎秒7cm以下)。南千島海峡における渦の構造は、底層では見られない。南千島海峡と太平洋の水交換は双方向に向かって行われている:色丹島より南では海峡に流入していて(毎秒1cm以下)、色丹より北では反対の動きになっている(毎秒1cm以下)。宗谷海流をモデルに入れる際には、流れの場と海底の地形の依存関係は保たれている。同時に、南千島海峡から太平洋への底層の海水の流出が一層はっきり現された。
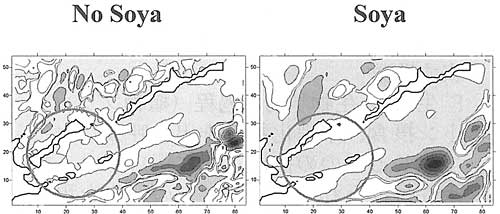
鉛直方向の流れのモデリングで、島々の太平洋側沿岸と南千島海峡は、幼生の着生に有利に働いていることがわかった、というのもそこでは流れの速さが大変遅いか、もしくは下に向かっているからである。宗谷海流があると、南千島海峡において海流が下方に向かう面積はより広くなり、同時に択捉島より南の部分では水はニュートラルな状態か、もしくは上昇している。
Field of surface flows (modeling time - 30 day)
Field of bottom flows (modeling time - 30 day)
Field of vertical flows (modeling time - 30 day)
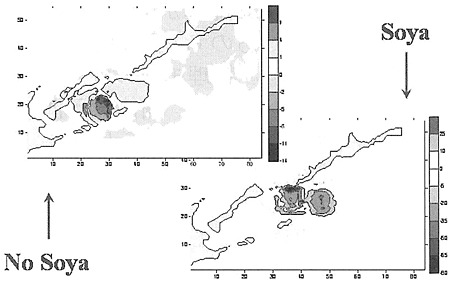
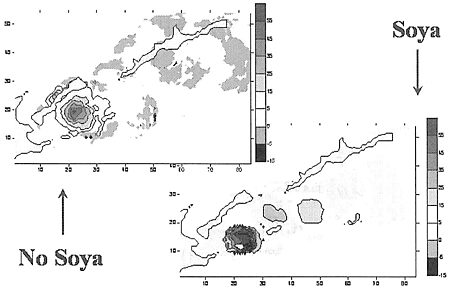
ハナサキガニの幼生の行動を分析するために四つの分布を考えた:表層、中層、底層、水柱の平均、である。表層で外部の流れがない場合、南千島海峡の北部における幼生の濃度が高まることが見られる。
国後島南沿岸のあたり、色丹・歯舞群島の島々の間と択捉島流があると、幼生の濃度の増大は、国後水道の出口部分でのみ見られる。
幼生の初期の分布の場所では、幼生の濃度の減少が見られ、そのことは、幼生がより低い層に分布していること、鉛直方向の速度がマイナスであることと一致する。
一番目のケースでは、南千島海峡の比較的大きくない海域で、中層に幼生が蓄積する場所と減少する場所があって、そのことから、鉛直方向の速度が変わりやすいことを指摘することができる。
第二のケースでは、全体図は本質的には変わらない。底層の濃度の分析によって、外部からの擾乱が無い場合、択捉島の幼生は消滅し、最大の濃度の斑点は、南千島海峡の中央部に移転する。底層の幼生の濃度は、実質上南千島海峡の全ての水域で上昇したことに注目しなくてはならない。
大体同じような変化の特徴が、宗谷海流がある場合にも見られる。濃度の増大するゾーンはもっと狭い面積になるが、色丹・歯舞群島にかかっており、その際幼生の数は本質的に増大する。
宗谷海流がない場合、水柱の幼生の濃度の平均の特徴は、底層における幼生の濃度の特徴と同じになり、水平方向の海流の弱さと水が常に沈降していることが指摘できる。宗谷海流があると、南千島海峡の南部におけるこれらの濃度の分布の図には、実際上マイナスの相関関係がみられる。この状態から、水平方向の平均的な流れと恒常的な水の沈降があることを見ることが出来る。
| Concentration of Paralithodes brevipes larvae in surface layer (k=1) |
| Concentration of Paralithodes brevipes larvae in middle layer (k=3) |
| Concentration of Paralithodes brevipes larvae in surface layer (k=5) |
| Average concentration of Paralithodes brevipes larvae |
結論
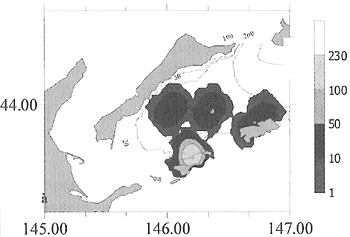
数値シミュレーションの結果、カニの幼生の最も可能性の高い着生場所は、色丹・歯舞群島の水域であることがわかった。同様に、国後島より南にも幼生が存在する可能性がある。カニの幼生が見られる確率は、得られた情報から、択捉島の水域では少ないことがわかった。
モデリングを行う際に、花咲カニの幼生の摂食や成長強度、天然状態における生存率などの情報がなかったために、シミュレーションは動物プランクトンの平均的パラメータを使った。それで得られた結果は、当然のことながら批判的にみる必要があり、確認が必要である。しかしながら私たちの考えでは、この数値実験から、生態−物理モデルをカニの幼生の分布と濃度の変化を予想するために利用できることは、明らかであり、個体数予測の基盤として使うこともできると言える。
この地域のモデルはさらに次のように発展させていく:
・潮流のモデリング
・カニ幼生の鉛直方向の移動を加える
・太平洋のここでは情報として必要ない大きな部分を除くために、モデル地図の転換を考えてみる
・風、気圧などを地図に入れ込む
・この地域についての追加的な情報を探し、追加する
| Real distribution of adult crab (A. Klitin, SakhNIRO, 2003) |
参考文献
1) Mellor, G. L. Users guide for a three-dimensional, primitive equation, numerical ocean model/G. L. Mellor//2004. -56 p.
2) Kawamiya, M. An ecological-physical coupled model applied to station Papa./M. Kawamiya, M. J. Kishi, Y. Yamanaka, N. Suginohara/ J. Oceanogr. -vol.51.-1995. -P. 635-664.
3) Oshima, Y. Evaluation of the nutrient budget in a seagrass bed. Ecological Modelling/Y. Oshima, M. J. Kishi, T. Sugimoto//vol. 115. -1999. -P.19-33.
4)レオーノフ A.V. 有機物形成物質の変化の水生物化学モデルとそのオホーツク海のエコシステムにおける一次生産物計算への適用/A.V.レオーノフ, V.V.サポージニコフ//オホーツク海エコシステム総合調査(V.V.サポージニコフ監修). −モスクワ:VNIRO出版−1997. -C.143-167.
5)ピシャリニク V.M サハリン島大陸棚海洋学地図/V.M. ピシャリニク, A.O. ボプコフ//ユジノサハリンスク:サハリン国立大学出版部, II部. -2002. -108c.
6)ベンツェリ M.V. 1990年冬および春の初めのオホーツク海の植物プランクトン./M.V.ベンツェリ//オホーツク海エコシステム総合調査(V.V.サポージニコフ監修). −モスクワ:VNIRO出版−1997. -C.205-209.
7)ナイデンコ S.V. 1990年代の南千島海域中心部のバイオマスの構成と動態、動物プランクトンの摂食/S.V.ナイデンコ//海洋生物学−2002. -T.28.-No.4.-C.272-278.
8)クリチン A.K. 1998年、1999年の南千島列島の漁獲対象カニの幼生の分布/A.K.クリチン//TINRO報告. -2002. -T.131. -C.266-283.
|