|
コブイシモ(Hydrolithon reinboldii)の単藻培養法
一木重夫
特定非営利活動法人
小笠原自然文化研究所
小笠原ホエールウォッチング協会
Method for uni−algal culture of Hydrolithon reinboldii(Rodophyta, Corallinales)
S. Ichiki
●要旨
紅藻の無節サンゴモの一種コブイシモ胞子を用いた単藻培養法を確立するため、胞子の生長に及ぼす抗生物質、振とう、水温、光条件の影響を調べた。その結果コブイシモ幼胞子体の培養には、抗生物質混液濃度が1ml/L、水温29℃、光量8μmol Photon s・m−2・s−1の環境条件が最適であることが明らかになった。また、静置培養が適当であると考えられた。
●緒言
紅藻の無節サンゴモはサンゴ礁域において死サンゴの塊同士を固着させる役割を果たしており、サンゴ礁形成に重要な役割を果たしている(Johansen 1981)。ハワイ諸島ワイキキのサンゴ礁におけるコブイシモ(Hydrolithon reinboldii)の被度はサンゴ礁全体の10.6%にも達していて、サンゴモ全体の50%を占めていることが知られている(Johansen 1981)。また最近、コブイシモの放出するアレロケミカルスが、サンゴ幼生の着生・変態を誘起することが明らかになっている(Morse et al. 1996)。この様に、コブイシモはサンゴ礁形成に重要な役割を果たしていることが明らかになってきている。しかし、無節サンゴモは藻体内に多毛類や原生動物等の小動物が生息していることが多く、また藻体表面にもコブイシモ以外の無節サンゴモが生育していることが珍しくないため、コブイシモだけを扱った生理学的な研究をする際に障害となることがある。そのため、単藻培養をして藻体をある一定の大きさまで生長させることが実験上必要になってくる。しかし、無節サンゴモは他の紅藻に比べて単藻培養が困難であり、無節サンゴモを扱って単藻培養を試みた研究報告は数少ない。そこで、本研究ではコブイシモ胞子を用いた単藻培養法を確立するため、胞子の生長に及ぼす抗生物質、振とう、水温、光条件の影響を明らかにすることを目的とした。
●材料と方法
コブイシモは1999年5月21日及び6月17日に沖縄県座間味村阿嘉島ヒズシで採集し1−3日後に水温21−24℃、1.92−9.6μmol Photons・m−2・s−1の光条件下で胞子を放出させ、放出後直ちにピペット洗浄法で胞子を単離した。単離した胞子を培養器(35mm/Non−treated Dish, Iwaki)に移し、水温21−24℃、24−32μmol Photons・m−2・s−1、光周期14L:10Dで2−5日間予備培養した。培養液は1/4PES(ビタミン無し:西澤一俊・千原光雄1979)に抗生物質混液(Tatewaki et al. 1989;表1)を1ml/Lの濃度で加えた滅菌海水を用いた(ただし、抗生物質混液の影響を調べる実験の予備培養では、抗生物質混液は加えなかった)。尚、以下に述べる実験においては、珪藻の繁殖を抑えるために、さらに0.25mg/L二酸化ゲルマニウムを加えた。
抗生物質の影響を調べる実験では抗生物質混液を0、0.5、1、2.5、5、10ml/Lの6条件下の濃度に設定し、インキュベータ(光照射式低温恒温器、Thomas科学機械)内で水温29℃(±1℃)、192μmol Photons・m−2・s−1条件下で培養した。振とうの影響を調べる実験では、胞子着底後に直ちに振とう器(Multi Shaker MMS、Tokyo Rikakikai Co. )を用いて100R.P.M. で浸とうさせ、水温24℃、24μmol Photons・m−2・s−1条件下で10日間培養した。尚、コントロールとして静置培養した培地も設定した。水温の影響を調べる実験では、水温21℃(±1℃)、25℃(±1℃)、29℃(±1℃)、192μmol Photons・m−2・s−1条件下に設定して12日間培養した。光の影響を調べる実験では、水温29℃で、1.6、8、16、32、64、128及び192μmol Photons・m−2・s−1の7条件下を設定して10日間培養した。各実験とも水換えは5日毎に行った。
表1. 抗生物質混液の組成(Tatewaki et al. 1989を改変)
| 内容物 |
量 |
| ポリミキシンB硫酸塩 |
3.16mg |
| ペニシリンGカリウム |
69.93mg |
| クロラムフェニコール |
1mg |
| 硫酸スプレプトマイシン |
250mg |
| 蒸留水 |
10ml |
|
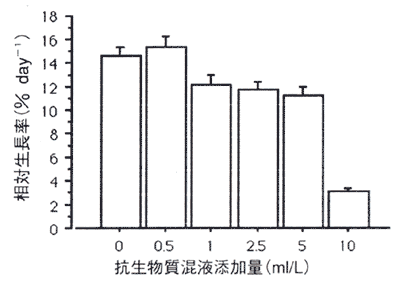
図1. 生長に及ぼす抗生物質混液の影響
値は平均値で、バーは標準誤差を示す(n=9-37)。
コブイシモ表面積は、培養開始時と培養終了後に顕微鏡下で長径を測定し、短径を長径×0.8と見積って、楕円に近似して算出した。そして、算出した面積から下の式を用いて培養期間の日間相対生長率(単位面積当たりの日間増加率を表す。単位は%)を求めた。
日間相対生長率=[100×{LN(培養後の面積)−LN(培養開始時の面積)}÷培養日数]
結果は、分散分析とScheffeテストを用いて検定した。
●結果
生長に及ぼす抗生物質混液の影響のグラフを図1に示す。抗生物質混液添加量が0ml/Lから5ml/Lの間で日間相対生長率は11.3−15.4%の範囲であり、実験終了時には0.07〜0.14mm2になっていた。添加量0.5ml/Lと5ml/Lの間に有意差があった(P=0.0397)。
添5加量10ml/Lの日間相対生長率は3.1%となり、他のすべての条件下と比べて有意に低かった(P<0.0001)。また、添加量10ml/Lで培養された藻体の色や形は他の培養条件の藻体と比べてほとんど差はなかった。尚、すべての条件下で枯死した藻体はほとんど見受けられなかった。
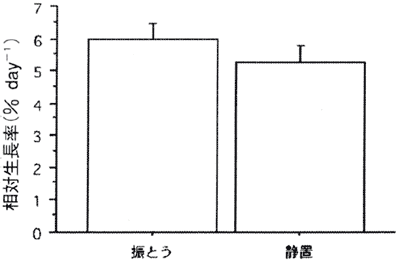
生長に及ぼす振とうの影響のグラフを図2に示す。振とう条件と静置条件の日間相対生長率はそれぞれ6.0%と5.3%で振とう条件の方が高かったが、有意差はなかった。
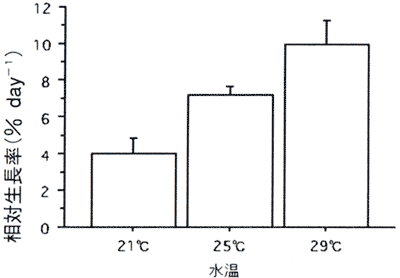
生長に及ぼす水温の影響のグラフを図3に示す。日間相対生長率は水温条件が高くなるほど高くなった。21℃条件で4.0%、29℃条件で9.9%となり両条件の間に有意差が認められた(p=0.004)。しかし、21℃条件と25℃条件、また25℃条件と29℃条件では有意差はなかった。
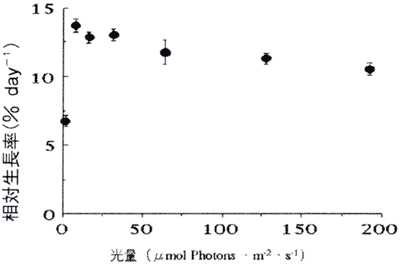
生長に及ぼす光の影響のグラフを図4に示す。1.6μmol Photons・m−2・s−1条件下で日間相対生長率は6.7%に過ぎなかったが、8μmol Photons・m−2・s−1条件下では約2倍の13.7%となっていた。結果を整理すると、8−192μmol Photons・m−2・s−1条件下での日間相対生長率は10.6−13.7%の範囲であり、1.6μmol Photons・m−2・s−1条件下と8−128μmol Photons・m−2・s−1条件下の間には有意差が認められた。
●考察
無節サンゴモは体の厚さが500μm以下を‘薄い'、500μm以上を‘厚い’藻体と分類できる(Steneck 1986)。無節サンゴモを単藻培養する際に、特に‘厚い’藻体の胞子は初期生長が遅いためか菌類やバクテリアに弱く、抗生物質を使用しないで胞子をある一定の大きさ(直径約1mm程度)まで培養することは非常に困難である(著者私見)。コブイシモの培養の場合、抗生物質混液の濃度が5ml/L以上で生長が阻害され、特に10ml/Lでは著しく生長が阻害された。このようなコブイシモの抗生物質混液に対する傾向は、他の無節サンゴモ(エゾイシゴロモとウミサビ)とほぼ同じであった(著者未発表)。本実験結果と他の無節サンゴモの培養で観察した菌類やバクテリアの発生状況(著者未発表)を考慮すると、菌類やバクテリアの大量発生を抑制し、かつコブイシモの生長にほとんど影響を与えないで培養することができる抗生物質混液の濃度は1ml/Lであると判断した。
図2. 生長に及ぼす振とうの影響
値は平均値で、バーは標準誤差を示す(n=15、18)。
著者の知る限り、これまでに無節サンゴモの生長に及ぼす振とうの影響に関する報告はない。本実験結果から振とうさせた方が若干生長が良かったが、有意差は認められなかった。本実験は水温24℃という最適水温よりも低い条件で実験したため、大きな差が現れなかったと考えられる。ただ、振とう機材を設置する手間と経費、振とうすることによって得られる生長への利点の大きさを考慮すると、コブイシモ胞子の初期生長段階においては静置培養で十分対応可能であると判断した。
無節サンゴモの生長に及ぼす水温の影響に関する研究は様々な種で報告されている。その大部分の研究報告では、その種が生育する海域の水温特性に適応した生長特性を有していることが明らかになっている。沖縄海域の水温は約21℃−29℃の幅で推移しており、コブイシモ幼胞子体の場合も沖縄海域の水温特性に適応した生長特性を有していることが明らかになり、コブイシモの初期生長段階では29℃が培養に適していると判断した。
図3. 生長に及ぼす水温の影響
値は平均値で、バーは標準誤差を示す(n=17−20)。
図4. 生長に及ぼす光量の影響
値は平均値で、バーは標準誤差を示す(n=13−22)。
無節サンゴモはその生育する光環境によって光特性が大きく異なる(Ichiki2001)。本研究では8μmol Photons・m−2・s−1で日間相対生長率がピークに達した。これは本種が主にサンゴの陰に生育しており、暗い光環境に適応した結果であると考えられる。北海道南西部に生育しているエゾイシゴロモは主に光が遮られることのない環境に生育しているが、その幼胞子体の相対生長率が飽和する光量は240μmol Photons・m−2・s−1であることが報告されている(Ichiki et al. 2000)。コブイシモの場合8μmol Photons・m−2・s−1以上の光量になると相対生長率が減少傾向になったが、明らかな強光生長阻害現象は認められなかった。これらのことからコブイシモの初期生長段階では特に強い光を照射する必要はなく、8μmol Photons・m−2・s−1程度であれば生長が制限されることなく培養できると判断した。
●謝辞
本研究を行うにあたり多大な協力を頂いた(財)熱帯海洋生態研究振興財団・阿嘉島臨海研究所の大森信所長及び岩尾研二研究員に深く感謝します。また、本研究をご指導頂いた北海道大学大学院の山本弘敏教授、安井肇助教授、水田浩之助手、東京水産大学の能登谷正浩教授、(財)海洋生物環境研究所の馬場将輔主任研究員に深く感謝します。原稿のご校閲を頂いた特定非営利法人小笠原自然文化研究所の稲葉 慎主任研究員に感謝の意を表します。
●参考文献
Ichiki, S., H. Mizuta and H. Yamamoto 2000. Effects of irradiance, water temperature and nutrients on the growth of sporelings of the crustose coralline alga Lithophyllum yessoense Foslie(Corallinales, Rhodophyceae). Physiological Research, 48:115−120.
Ichiki, S. 2001. Study on propagational mechanisms of Lithophyllum yessoense in southwestern Hokkaido. PhD thesis, Hokkaido Univ.
Johansen, H. W. 1981. Coralline Algae, A First Synthesis. CRC Press, Inc., Boca Raton, Florida. 233PP.
Morse, A. N. C., K. Iwao, M. Baba, K. Shimoike, T. Hayashibaa and M. Omori 1996. An ancient chemosensory mechanism brings new life to coral reefs. Biol. Bull., 191:149−154.
西澤一俊・千原光雄 編 1979. 藻類研究法.共立出版、東京.754pp.
Steneck, R. S. 1986. The ecology of coralline algal crusts:convergent patterns and adaptative strategies. Annual Review of Ecologh and Systematics, 17:273−303.
Tatewaki, M., W. Xiao−Yang and I. Wakana 1989. A simple method of red seaweed axenic culture by sporewashing. Japanese Journal of Phychology, 37:150−152.
徳田 廣・大野正夫・小河久朗 1989. 海藻資源養殖学.緑書房、東京。354pp.
|