|
千葉県館山湾坂田地先に生息する造礁
サンゴ類の分布、生息環境および有性生殖
萩原良太
東京水産大学大学院
Distribution, environmental conditions and sexual reproduction of the zooxanthellae corals at the Banda area of Tateyama Bay, Chiba, Japan
R. Hagiwara
●はじめに
房総半島の館山湾は、黒潮が本州から離岸して東方へ向かう北緯35°というライン上にあって、そこはイシサンゴ類を含む多くの亜熱帯性海産動物の分布北限となっている(藤岡1994)。日本の造礁サンゴ類はその分布範囲からトカラ海峡以南にのみ出現する沖縄型と、伊豆半島や房総半島まで見られる南日本沿岸型に分けられるという考えがあり(福田ほか1991、野島ほか1992)、館山湾に生息する種は沖縄などの亜熱帯サンゴ礁域では全く出現しないか、また出現してもその数は少ない。
館山市の沼と呼ばれる海抜15〜20mの所には沼サンゴ層とよばれる約5000〜6000年前の化石サンゴ層がみられ、地質学的にも重要な場所として知られている。そこでの化石サンゴの研究では、千葉県地学教育研究会(1963)が74種、江ロほか(1973)が約100種を報告している。最近ではVeron(1992b)が55種の化石を分類し、また過去の知見と合わせて当時は約70種が生息していたと考えている。そして沼サンゴ層での出現種のうち、現在生息している19種を含め、館山湾で24種の分布を報告した。
近年、館山湾では8月下旬にAcropora tumidaとFavida sp. の産卵が確認されて(三瓶私信)、マスコミでも取り上げられた。またAlveopora japonicaでも有性生殖が報告されている(Harii et al. 2001)。これらによって分布北限域でも体内に共生藻をもつ造礁サンゴ類は有性生殖により、種を維持・拡散している可能性が出てきた。本報は、館山湾坂田地先での造礁サンゴ類の分布と生息環境を調査し、成長速度と有性生殖を測定・観察して、分布北限域での造礁サンゴ類の生態を明らかにしようとしたものである。
●方法
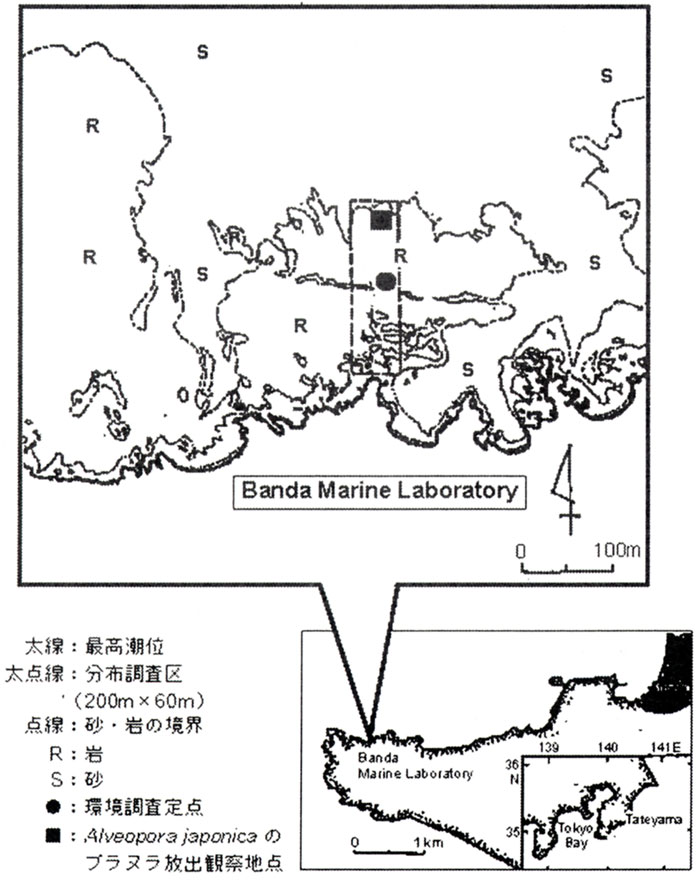
造礁サンゴ類がみられる坂田地先の水深5m付近に観測定点を設定し(図1)、海域の水温、塩分、懸濁物量(沿岸環境調査マニュアルII. 日本海洋学会編1990による定法)を1996年12月から1997年11月まで毎月1回、水中照度(水中照度計:ALK−IIアレック電子株式会社製による)を1997年7月24日、8月13、24日、1998年1月25、26日に測定した。また、1994年2月以降の水深5mの水温・塩分の変化を東京水産大学坂田水産実験所の自記記録計のデータから得た。
| (拡大画面:101KB) |
 |
図1. 坂田実験実習場における調査区の位置
表1. 分布調査区内に出現した造礁サンゴ類と群体数およびその生息水深
| 種名 |
群体数 |
生息水深(m) |
| Family Astrocoeniidae(ムカシサンゴ科) |
|
|
| Stylocoeniella guentheri Bassett-Smith, 1890(ムカシサンゴ) |
65 |
3.0-7.2 |
| Family Poritidae(ハマサンゴ科) |
|
|
| Alveopora japonica Eguchi, 1968(ニホンアワサンゴ) |
227 |
4.5-8.3 |
| Porites heronensis Veron, 1985(フタマタハマサンゴ) |
6 |
3.6-6.9 |
| Family Siderastreidae(ヤスリサンゴ科) |
|
|
| Psammocora profundacella Gardiner, 1898(アミメサンゴ) |
11 |
3.1-7.2 |
| Psammocora superficialis Gardiner, 1898(ベルベットサンゴ) |
5 |
5.7-5.9 |
| Family Merulinidae(サザナミサンゴ科) |
|
|
| Hydnophora pilosa Veron, 1985(イボサンゴ) |
1 |
6.6 |
| Family Faviidae(キクメイシ科) |
|
|
| Favia favus Forskal, 1775(スボミキクメイシ) |
1 |
6.0 |
| Favia speciosa Dana, 1846(キクメイシ) |
1 |
6.5 |
| Goniastrea deformis Veron, 1990(ミダレカメノコキクメイシ) |
6 |
5.8-7.0 |
| Oulastrea crispata(Lamarck, 1816)(キクメイシモドキ) |
22 |
4.8-8.1 |
| Plesiastrea versipora(Lamarck, 1816)(コマルキクメイシ) |
2 |
6.2-6.4 |
|
造礁サンゴ類の群体分布と周辺環境の調査は、1997年7月16日〜11月22日の間に、2人組でスキューバ潜水および素潜りで行い、約60m×200m(水深0〜8m)の調査区を設定し、その範囲内に出現した造礁サンゴ類を同定し、それぞれの群体の長径・長径と直角に交わる最大径・高さなどを測定した。同時に、底質とサンゴ群体周辺にみられる藻類・ベントス類も調べた。調査区内に出現した造礁サンゴ周辺すべてを50cm×50cmのコドラートで被い、その中をデジタルビデオで撮影し、家庭用の大型モニター画像に再生して調査の正確性を期した。造礁サンゴ類の種の同定には西平(1991)、西平・Veron(1995)、Veron(1985)などを用いた。
館山湾の造礁サンゴの成長速度は、1997年9月〜同年12月の約3か月間、Acropora tumida2群体と、Stylocoeniella guentheri4群体、およびPsammocora profundacella2群体について調べた。この内A.tumidaは同湾、沖の島地先に分布したものである。群体サイズ(長径)はA.tumidaが約25cm、S.guentheriが約6cm、P.profundacellaが約60〜100cmであった。それらを水温26℃、12時間明期12時間暗期の恒温飼育水槽中で1週間以上馴致させた後、A.tumidaは枝の先端部を折って、4cm×4cmの塩ビ板にアロンアルファで固定し、S.guentheriとP.profundacellaは、ペンチで砕いて約1.5cm四方の小片にして、エポキシ水中ボンドで4cm×4cmの塩ビ板に接着した。そして、水温14℃区(館山湾の3月の平均水温)、25℃区(同、8月の平均水温)、コントロール(流水)区の3実験区を恒温飼育水槽に設置し、サンゴ固着基盤をそれぞれ6個、4個、6個ずつ入れた。光条件は12時間明12時間暗とし、明期の照度は7000luxに設定した。別に、採集現場近くにも野外実験区を設け、同じサンゴ固着基盤を同数設置した。これらは1ヶ月毎に、ノギスを用いて骨格の接着面からの「高さ」を0.005mm単位まで計測した。また、水槽中で固着基盤をのせたシャーレを吊り下げ、電子天秤(メトラーAT200)を用いて、水中重量を測定した。また、3ヵ月後に2次元的な成長を確認するための写真撮影を行った。そしてそれらの結果を基に、年間成長速度を推定した。
| (拡大画面:101KB) |
 |
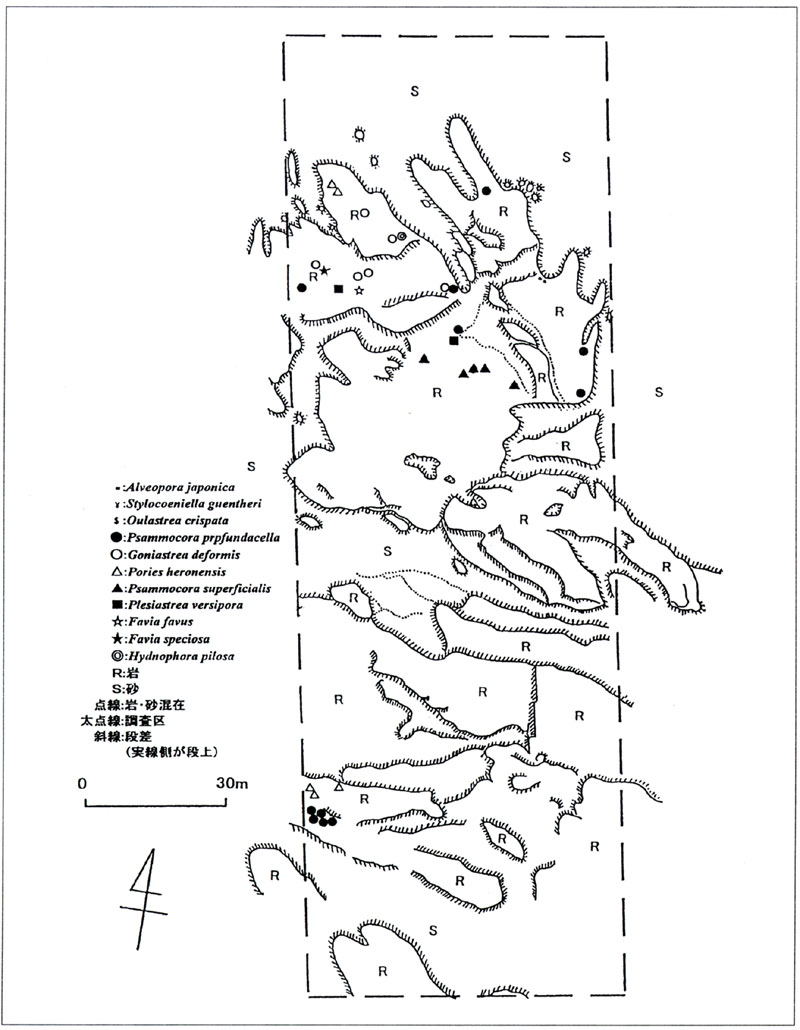
図2. 坂田地先における造礁サンゴ類の分布
| 表2. |
調査区内の造礁サンゴ類の水深別単位面積当たり群体数(×0.1/100m2)および多様度示数 |
| 水深(m) |
3.0−3.9 |
4.0−4.9 |
5.0−5.9 |
6.0−6.9 |
7.0−7.9 |
8.0−8.9 |
| 各水深の基底面積(m2) |
1089 |
978 |
1845 |
1797 |
2112 |
426 |
| Alveopora japonica |
0.0 |
3.1 |
1.1 |
22.8 |
66.3 |
98.6 |
| Stylocoeniella guentheri |
8.3 |
15.3 |
4.9 |
14.5 |
2.8 |
0.0 |
| Oulastrea crispata |
0.0 |
1.0 |
0.5 |
2.2 |
5.7 |
9.4 |
| Psammocora profundacella |
4.6 |
0.0 |
1.1 |
1.7 |
0.5 |
0.0 |
| Goniaastrea deformis |
0.0 |
0.0 |
1.1 |
1.7 |
0.5 |
0.0 |
| Porites heronensis |
1.8 |
2.0 |
0.0 |
1.1 |
0.0 |
0.0 |
| Psammocora superficialis |
0.0 |
0.0 |
2.2 |
0.0 |
0.0 |
0.0 |
| Plesiastrea versipora |
0.0 |
0.0 |
0.0 |
1.1 |
0.0 |
0.0 |
| Favia favus |
0.0 |
0.0 |
0.0 |
0.6 |
0.0 |
0.0 |
| Favia speciosa |
0.0 |
0.0 |
0.0 |
0.6 |
0.0 |
0.0 |
| Hydnophora pilosa |
0.0 |
O.0 |
0.0 |
0.6 |
0.0 |
0.0 |
| 多様度H' |
0.089 |
0.116 |
0.081 |
0.235 |
0.213 |
0.209 |
|
Acropora tumidaとAlveopora japonicaの産卵についてはそれぞれ沖の島地先と坂田地先に見られた群体数個について確認し、A. japonicaについては放出されたプラヌラを採集して水槽中で行動を観察した。
●結果
1994年2月から1997年7月までの4年間の平均水温(水深5m)は、最高が8月の23.8℃、最低が3月の14.3℃で、12月〜5月の6ヶ月間は、18℃以下であった。塩分は年間を通じておよそ33.0〜35.0PSUで安定しており、年間平均は34.3PSUであった。懸濁物量は、0.50mg/l(1997年1月)〜2.88mg/l(1997年4月)で、平均1.27mg/lであった。水中照度は、夏季(1997年7〜8月)11,000〜30,000lux(平均18,677lux)、冬期(1998年1月)7,800〜26,000lux(平均6,900lux)であった。因みに、懸濁物量の年平均値は沖縄県阿嘉島(林原1995)のそれより、2.1〜4.0倍大きな値であった。懸濁物量は1月に最低値を記録したにもかかわらず、水深5mの照度は夏期と冬期で近似していた。これは、冬期に大型褐藻類を中心とした藻類が急速に成長し、水中の光を遮った結果と考えられる。
造礁サンゴはこれまで館山湾で記録のなかったStylocoeniella guentheriとFavia favusを含む11種347群体の分布が確認された(表1、2、図2)。各種分布について述べると、Alveopora japonicaは最も多く出現し、砂地と岩場の境目の岩盤斜面によくみられ、多いところでは30群体/m2ほどのパッチ状群落がみられた。群体の長径は0.1〜7.4cmであった。Stylocoeniella guentheriもパッチ状に長径10cmに満たない小さな群体が分布しており、全出現種中もっとも浅い水深3mにも生息していた。Oulastrea crispataは周りがほとんど砂に囲まれているような場所でよくみられ、周辺には石灰紅藻類・アラメ・マメダワラが多かった。一般に本種は長径10cm以内の群体が多いとされている(西平・Veron 1995)が、坂田地先では18.3cmのものが確認された。Psammocora profundacellaは水深3.1〜7.2mに長径18.0〜107.0cmの群体がみられ、サイズも生息水深もかなりばらついた。また、3m付近にみられた群体は、その形状から、同一群体が部分死して分かれたものと思われた。Goniastrea deformisは、水深5.8〜7.0mの、割合平らな岩盤上にみられ、群体サイズは長径で18.0〜95.0cmであった。Porites heronensis水深3.6〜6.9mでパッチ状に分布していた。群体サイズは長径で23.8〜95.0cmであった。Psammocora superficialisは水深5.7〜5.9mの狭い範囲にみられ、長径37.0〜105.0cmであった。Plesiastrea versiporaは水深6.2mと6.4mの平らな岩盤上に2群体が確認された。群体の長径はそれぞれ148.0cm、68.0cmであった。Favia favus, F. speciosa、およびHydnophora pilosaは、それぞれ水深6.0m、6.5m、6.6mに1群体ずつ出現した。Favia favusは若干傾斜のある岩に付着していたが、後の2種はほぼ平らな岩盤上にみられた。いずれのサンゴ類でも群体の死亡部分には石灰紅藻類や紅藻類や緑藻類がよくみられ、ケヤリムシ類が穿孔している群体もみられた。
Shannon−Weaverの情報指数H’で表した種の多様度示数は、水深6.0〜6.9mの間で最も高く、次いで7.0〜7.9mであった。水深3m以浅ではサンゴ類はみられなかった(表2)。別に、群体の面積をGrigg & Matagos(1974)の方法(S=(R・r)/4π但しSは群体の面積、Rは長径、rは長径と直角に交わる最大径)で求め、水深別の基底の面積と群体の面積から、各水深毎の種別の占有率を求めた結果、占有率は水深6.0〜6.9mの間で最も高く、約0.3%で、5.0〜5.9m間の約0.1%がこれに次いだ。また、全調査区内(60m×200m)の造礁サンゴ類の占有面積は0.08%であった。
Stylocoeniella guentheriとPsammocora profundacellaおよびAcropora tumidaの成長速度を表3、4に示す。実験期間中に死んだ群体がかなりあったので、測定結果が必ずしも正確とは言えないが、成長速度は14℃区より25℃区の方が大きく、現場実験区よりコントロール区がいく分大きいようであった。
Acropora tumidaの産卵は1996年9月4、5、7、8日の午後8時20〜40分と、1997年8月19、20、22、23、25〜27、30〜9月2日、7〜9日の午後7時53分〜9時20分に確認された。産卵は全て日没から約2時間10分後に開始されている。1996年は2群体で、それぞれの一部分ずつが産卵したので、群体としては4日間連続して産卵したが、同種の他群体との産卵の同調は確認されなかった。1997年は2群体あるいは3群体が同調して産卵し、同一群体で6〜11日間に渉って分割産卵をした。産卵と月齢周期の関係は見られなかった。またこの間、他の種の産卵は観察されなかった。
一方、Alveopora japonicaは、1群体のみがプラヌラ幼生を放出した。1997年10月24日の午前9時20分〜10時37分の間に17個体、25日は午前9時25分〜10時41分の間に32個体、28日は午前10時8分〜11時2分の間に20個体が放出された。全ての幼生は、こん棒状の形態(長径1.83±0.28mm(n=19))であった。プラヌラを飼育瓶に移し、着生のための基盤を入れたところ、計13個体が着生した。着生したプラヌラは正常に発生し、約2週間で変態を完了した。
表3. 3種の造礁サンゴの89日間平均成長(高さ、mm±1SD)
( )内の数値は標本群体数を示す。
| |
14℃ |
25℃ |
コントロール
(流水) |
野外 |
| Stylocoeniella guentheri |
- |
0.35(1) |
0.05(1) |
坂田 0.45(1) |
| Psammocora profundacella |
0.03±0.06(6) |
0.95±:0.53(6) |
0.69±0.28(6) |
坂田 0.43±0.10(6) |
| Acropora tumida |
0.34±0.36(6) |
4.75±:6.30(6) |
4.67±4.64(6) |
沖の島 0.10±0.20(4) |
|
表4. 3種の造礁サンゴの日間平均増加量(mg/日±1SD)
( )内の数値は標本群体数を示す。
| |
14℃ |
25℃ |
コントロール
(流水) |
野外 |
| Stylocoeniella guentheri |
0.702(1) |
0.478(1) |
0.875(1) |
坂田 0.795±0.105(3) |
| Psammocora profundacella |
0.421±0.180(6) |
4.176±1.922(6) |
2.180±1.119(6) |
坂田 2.191±0.566(6) |
| Acropora tumida |
0.380±0.258(5) |
9.049±2.097(6) |
6.502±3.836(6) |
沖の島 1.871±0.823(5) |
|
|